一名生态学家的数学探索
时间:2023-04-27 01:14:09 来源:科普之家 作者:返朴 栏目:头条 阅读:114
Eugene Wigner的名篇《数学在自然科学中不可思议的有效性》,单从题目来看即蕴含着两重思想:1. 数学对于理解自然的必要性;2. 自然现象在抽离出研究对象以丰富数学思想方面的重要性。混沌理论的思想起源于不同领域的不同专家对于不同自然现象的数学描述的孜孜以求,最终殊途同归,抵达了统一的认识。本文旨在讲述生态学家在用逻辑斯蒂映射描述种群动力学行为时发现的倍周期分叉的数学特征。
撰文 | 丁玖(美国南密西西比大学数学系教授)
我在之前的《返朴》文章《这么说迭代,你一定能懂》(以下简称为《迭代》)中介绍了最基本的函数迭代现象,借多项式这一简单道具,合用几何法和分析法,展示了吸引或排斥的不动点或周期为二的周期点。有了这些基础后,我们可以继续行走在迭代的大道上,一路采撷五彩缤纷的朵朵鲜花。
这篇文章为读者奉献的是一簇簇特别绚丽的玫瑰之花,它们盛开在连绵不断大枝分叉成小枝的迭代大树上,它们也为组成“迭代春游团”的团员历经一场从有序到无序之旅,特地建造了一座走进混沌世界之前的缓冲装置,让他们在猛见混沌怪兽而发出一声惊叫前吸足气、定定神。这整片的花朵开在带有一个参数的一系列二次多项式的花园里。
然而,让“倍周期分叉”红玫瑰花朵光荣绽放的营养素却来自于生物科学的一个宏观分支——生态学,它也被称为种群动力学,为玫瑰树施肥的辛勤园工是后来担任过英国政府首席科学家并被伊丽莎白女王封爵的罗伯特·梅 (Robert May,1936-2020) 。
梅男爵生于澳大利亚大城悉尼,在悉尼大学本科毕业时才二十岁,读的是化学工程与理论物理。三年后,他在本校获得理论物理博士学位,接下来去哈佛大学做了两年博士后,研究应用数学。重返母校后,他回归本行,以高级讲师的职称开始教书,一路升到理论物理学正教授。1971年,因为又“心血来潮”地对生物学着了迷,他再度赴美,到普林斯顿高等研究院待了一年,并与普林斯顿大学的生物学家们交上朋友。两年后,他成为这所名校的动物学讲座教授。
正是在这里,他成了一位理论生态学家,并在基于函数迭代的创造性种群动力学研究中出了大名。这门学科从属于生物学中的一个分支“群落生态学”,探索的是共同居住在同一个地理范围或区域内的两种或更多种不同生物种群之间的相互依赖和制约关系,以及生物种群数目的涨落和生命的盛衰,目标是寻找这些千变万化的数量背后的规律、自然群落的复杂性和稳定性之间的关系,以及能够解释变化规律的数学刻画。
像一切其他自然科学家和工程技术人员,生态学家们自然也需要数学的帮助。然而,在从事像菲尔兹奖获得者斯梅尔 (Stephen Smale,1930-) 那样用深奥的现代数学研究动力系统的几何或拓扑学家眼里,梅闻名于世的研究所用到的数学似乎非常简单。事实上,他最为天下所知的工作只用到二次多项式,只不过其表达式中多了一个参数而已。然而这族函数的无穷多个弯曲度不一的抛物线图象,将他写出的科学论文像乔丹手中的篮球被一举抛进了《自然》和《科学》的期刊球篮里。

顺便一提,早在1947年,分别为波兰裔和匈牙利裔的美国数学家乌拉姆 (Stanislaw Ulam,1909-1984) 和冯·诺伊曼 (John von Neumann,1903-1957) 就对逻辑斯蒂映射族中μ = 4时这根最长的抛物线做出了精准的统计学研究,给出了对几乎所有初始点都一样的迭代点轨道最终分布规律,这将是我未来一篇关于遍历理论的科普文章中的一朵鲜花。
这个著名的数学模型符合生态学家对种群数量问题的直觉,就连大众也不难理解。比如,在弱肉强食的非洲大草原,食草动物斑马是弱者,而食肉动物狮子是强者,二者的种群数量相互制约,弱者如果被强者吃得太多,数目就会急剧减少,强者反过来也会面临生存危机。所以,经验告诉我们,如果种群数比较小,就会上升很快;而数目适中时会导致增长速度趋近于零,种群数量很大时则急剧下降。这是生态学家从事研究时遵循的一条基本假设,它也反映在上述模型中:如果x上涨,那么1-x就会下跌,反之,如果x减小,1-x就增加,故而它们的乘积就制约着种群数量的变化。看来,这种函数关系确实可以在一定程度上反映生存环境下种群数量变化的规律。
在梅教授进入这个领域之前,早期生态学家们普遍认同英国理论进化生物学家和遗传学家史密斯 (John Smith,1920-2004) 在其经典著作《生物学中的数学思想》(Mathematical Ideas in Biology) 中所主张的观点:种群数往往近似为常数。他们或多或少都相信,在如上的逻辑斯蒂映射中,无论初始的种群数有多大或多小,几年后数目就会稳定在一个固定的数。同时,他们纷纷认为,只有“稳定解”才吸引人。如果看到不肯稳定下来的种群数目,就会不假思索地认定是计算工具的误差在捣乱。
这种未经科学实验证实的信条已经在众多科学家的脑海里根深蒂固,因此他们极少肯坐下来对具体模型做一番数学分析的功课,更不要说无比耐心地从事枯燥无味的重复迭代运算。恰恰十年前的四月,哈佛大学已退休的一位著名生物学家威尔逊 (Edward Wilson,1929-2021) 在《华尔街日报》上撰文《伟大的科学家≠擅长数学》,试图用自身经历论证:“当今世界上许多最成功的科学家在数学上不过是半文盲”,该文以及四天后一名伯克利数学教授的反驳文章《不要听爱德华·威尔逊》在当年七月被《美国数学会会刊》转载后,引起不少数学家与科学家的热烈讨论。
作为身跨多个领域的科学家,梅教授显然不认同传统生物学家轻视数学的上述代表性观点,在普林斯顿大学,他开始了对逻辑斯蒂映射的数值迭代试验。正是集应用数学家和生态学家于一身的梅教授将数学耐心地施加于逻辑斯蒂映射族,通过亲手进行迭代计算,他在种群动力学的田野上看到了令他惊奇不已的一派新景象。
为了叙述的方便起见,在下面我们假定以年为时间段,即若x表示本年的相对种群数,则μx(1-x)表示下年的相对种群数。
梅逐步增加繁殖率μ,想把种群数演变的最终趋势与这个重要参数之间的关系搞个水落石出。很快他就发现,当μ不超过3这个数时,一切都很正常。比如说,如果参数μ不大于1,那么无论起先是什么样的种群数,最迟在第二年后,种群的数目就会逐年下降,最终将走向消亡。然而当μ在大于1但不大于3时,不管开始的种群数为几,一年年迭代之后的种群数会逐渐地稳定下来,最终将趋向于某一个固定的数。这个固定数随着参数的增加而增加,例如,当μ取值为2.7时,最终的种群个数会固定在0.6296,而当μ = 3时,终极种群数则增加到0.6667。
梅继续增加参数的值。
当μ大于3但不大于近似值约为3.45的一个精确值1 + √6时,他发现了新现象:种群数不再最终趋向于一个固定的数,而是一年年地交替升降,最后在两个不同的固定数附近之间不停地来回跳动,最终趋向于这两个周期点组成的一个周期-2轨道,而与迭代初始所选的种群个数无关。
如果让参数值比3.45再大那么一点点,直到差不多3.54时,种群数则每过四年才呈现出有规律性的涨落,最终在四个固定数附近之间周而复始地跳来跳去,最终趋向于这四个周期点组成的一个周期-4轨道,而无论种群的初始数有多大。这样一来,种群数的两年周期现象就加倍成四年周期现象。
类似地,当参数值超过3.54一个微小的数后,四年周期将跳到八年周期。然后,随着参数值不断细微增加,十六年周期、三十二年周期……,等等等等直到无穷,依次登场。然后,在μ的一个新的取值范围内,又有了周期不是2的次方的新的周期加倍现象。而当μ再增加到一定程度后,最终的种群数量则看不到周期性,出现了某种“乱七八糟”的迹象。
我们将上面几段用数学语言总结一下。

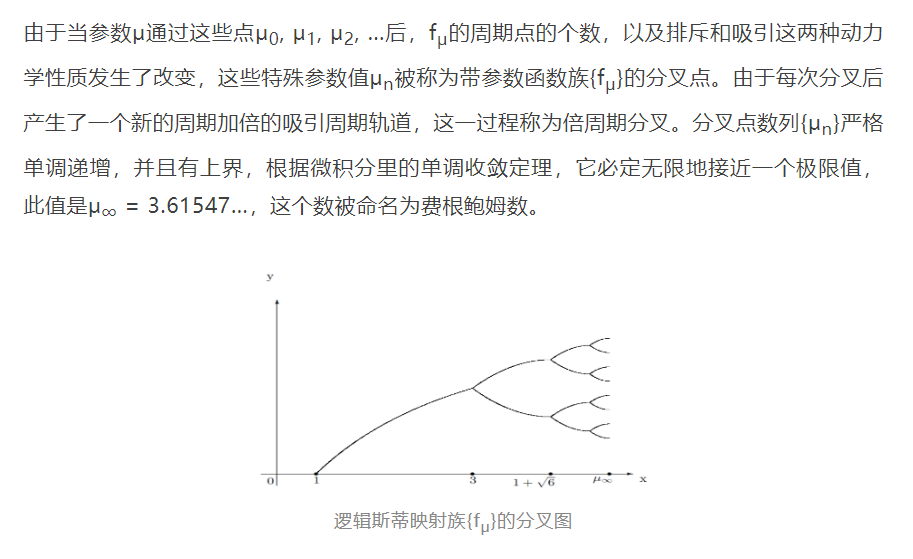
这个极限值得名如此,是因为美国数学物理学家米切尔·费根鲍姆(Mitchell Feigenbaum,1944-2019)。
费根鲍姆于1970年在麻省理工学院获得粒子物理博士学位,然后在康奈尔大学和弗吉尼亚理工学院暨州立大学各待了两年,这四年中他只发表了一篇论文,但积累了广泛的知识基础,可谓厚积薄发。事实上,他一生中独立完成或与人合作的科学论文也只有二十七篇。
他1974年被美国洛斯阿拉莫斯国家实验室招聘过去后,为以自己的方式深刻理解湍流中大小尺度之间的自相似性,并找到问题的本质,他全然不顾能不能尽快挤出文章,而是一天工作“二十五”个小时,靠他摆弄计算器的双手和计算间隙不停思考的大脑,发现了新的普适常数。他在理论部的顶头上司后来评价:“费根鲍姆具有正确的背景,他在正确的时候做了正确的事情,而且做得很出色。他不是做局部的事情,而是把整个事情弄清楚了。”


这是什么原因呢?
1974年,梅教授应马里兰大学数学系“生物数学系列演讲”之邀,报告了他对逻辑斯蒂映射数值迭代时发现的奇怪现象。演讲之后,把他请来的一位教授送他去机场。路上,那位名叫詹姆斯·约克 (James Yorke,1941-) 的教授递给梅一篇文章,一阅之下,梅教授大吃一惊:这篇文章里的一个定理解除了他的困惑。
这篇当时还只是初稿的文章后来成为混沌史上最著名的数学论文之一,如今它的题目在中文世界也有了固定翻译:“周期三则意味着混沌”。对于这篇已被学界引用超过5650次的区区八页数学论文的初等诠释,将是我不久后一篇数学科普文章的内容。
关于“倍周期分叉”数学现象的生态学故事算是大致讲完了,然而,光知道“科学人物的发现轶事”而缺乏对科学原理的基本认识,就如同美国理论物理学家费曼 (Richard Feynman,1918-1988) 经常回忆的他父亲在他还是孩提时特地忠告儿子的一句话“你如果对一只鸟只知道它的名字,而对它的习性却一无所知,那么你对那只鸟的了解几乎为零”所表达的真理。费曼认为父亲这种简单而有智慧的观点影响了自己的科学生涯一辈子,让他很早就懂得了仅仅知道事物的名称和充分了解事物的本质这一根本区别。因此,我在本文的最后一部分转向数学,主要用浅显易懂的初等数学语言解释上文关于逻辑斯蒂映射族fμ的三个倍周期分叉性质中,为何当繁殖率μ越过3这个门坎后一下子冒出一个周期-2轨道,并且该轨道在μ值超过1 + √6前都是吸引的。



我在《迭代》中讲过,线性函数ax + b当|a| < 1时,其唯一的不动点b/(1-a)是吸引的,但当|a| > 1时,则b/(1-a)是排斥的不动点。这里的系数a也是对应的直线图象的斜率。对于任意一个可微函数y = f(x),它在一点x的导数值f’(x)就是函数的图象在点(x, f(x))处的切线的斜率。非线性函数的图象曲线虽然是弯曲的,然而在切点的附近,曲线和切线看上去几乎一样,这个观察是为何几百年来微积分大获全胜的基本道理。既然不动点的“吸引性”或“排斥性”像“函数的导数”概念一样是个“局部性质”,我们凭什么不能借用在不动点处相切于函数图象的简单直线来代替复杂曲线制作数学工具,判断函数f的不动点x*是否具有“吸引力”或“排斥力”呢?这个思想至少能帮助我们“想出”一个简单易行的判别法,即如果f的不动点x*满足|f’(x*)| < 1,那么x*一定是吸引的;如果|f’(x*)| > 1,那么x*一定是排斥的。这个想法是千真万确的。
为什么呢?画一条在与对角线y = x相交点附近走向比较平坦的函数曲线,用“图象迭代法”很容易就能几何地证明上述断言。但下面的“分析论证”更加令人信服:假设f(x*) = x*及|f’(x*)| < 1,由于导数f’(x*)是函数值差f(x) – f(x*)与自变量值差x – x*的比当x趋于x*时的极限,我们有理由相信当x在x*附近时,|f(x) – x*| ≤ δ |x – x*|,其中正数δ只比|f’(x*)|大了那么一点点,但还是像|f’(x*)|那样小于1(比如,可以取δ为|f’(x*)|和1的算术平均,即δ = (|f’(x*)| + 1)/2)。

用上面的思想同样可以证明,如果f的不动点x*满足|f’(x*)| > 1,那么x*是排斥的,即它排斥了其附近的不等于它的每一点x:|f(x) – x*| ≥ Δ |x – x*|,其中Δ为某个大于1的数。
读者自然想到当|f’(x*)| = 1时结论如何。没有办法,没有一般性结论,只能是“具体问题具体分析”;就像人生一样,数学也有遗憾。学过高等数学中的无穷级数理论的读者定会记得这个事实:任给一个关于级数收敛与否的判别法,总能找到级数,对它而言该判别法失效。所以,世间没有“万能钥匙”;当我看到一些出版社诸如“世界公认最牛学习方法!”的促销广告词时,就免不了呵呵一笑,如同见到发明出永动机的新闻一样。


历史上首次由生态学家梅博士数值模拟的这类单参数函数族的分叉规律,在动力系统领域被形象地称为干草叉型的,因为上面的分叉图如果用虚线再画上那些排斥的周期点,就像是一个个农村里常用的工具“干草叉”,它与“倍周期分叉”不可分离。不过还有一种分叉现象,称为“切线型”的,它来自于在参数跨过一个门槛的前后,函数的图象历经一次与对角线y = x相交两点,相切于一点,到彼此分离的“合久必分”过程,因而不动点的个数从二减少到一再到零。它的一个典型例子是指数函数族{μex},其中参数μ > 0。当μ = 1/e时,函数图象与对角线相切于不动点1,当μ < 1/e时,图象与对角线相交于两点,而当μ > 1/e时函数图象与对角线绝不相交。作为读完本文后的自我测验,我邀请读者用“图象迭代法”对这族指数函数就0 < μ < 1/e,μ = 1/e及μ > 1/e这三种情形, 预测一下对应于所有初始点的迭代点轨道的最终走向。
到目前为止,我们的函数迭代仅仅碰到周期为2的非负整数幂次的周期点,包括不动点。在全然有序的自然数中,紧跟着2的是3,这可是一个不同凡响的自然数,它将在函数迭代的大海里掀起巨浪,让航行中大船的未来航线无法预见!
致谢:作者感谢学者杨运洋阅读初稿并提出修改建议。
出品:科普中国

特 别 提 示
1. 进入『返朴』微信公众号底部菜单“精品专栏“,可查阅不同主题系列科普文章。
2. 『返朴』提供按月检索文章功能。关注公众号,回复四位数组成的年份+月份,如“1903”,可获取2019年3月的文章索引,以此类推。
版权说明:欢迎个人转发,任何形式的媒体或机构未经授权,不得转载和摘编。转载授权请在「返朴」微信公众号内联系后台。
本文链接:https://www.bjjcc.cn/kepu/76648.html,文章来源:科普之家,作者:返朴,版权归作者所有,如需转载请注明来源和作者,否则将追究法律责任!